地球上的绝大多数植物都能够自立更生,从土壤中吸取养分,依靠自身光合作用获取生长能量,它们可统称为“自养型”。然而,在被子植物中,进化出了至少12类(约5000个物种)“好吃懒做”的寄生植物,它们寄生生在其它植物上,并从中偷取营养,有一些还保留了光合作用能力(被称作半寄生植物),另外有一些完全丧失了光合作用能力(被称作全寄生植物),蛇菰(gū)科植物就是全寄生植物的典型代表之一。
如果在森林中的某个角落偶然遇到蛇菰科植物,你很可能会将它们误认为是蘑菇。与蘑菇相似之处在于,蛇菰科植物也是异养的。但是蘑菇属于真菌类,而蛇菰是一种高等植物。蛇菰的形态经历了严重的退化,缺乏正常的根、茎和叶组织,主要的营养器官是根茎,花序从根茎中伸出,而根茎的另一端与寄主植物的根相连。由于其独特的生活方式,关于它的生长和发育,我们了解甚少:种子萌发后,首先产生吸器,吸器会占据寄主植物的根尖,导致寄主根停止生长,随后蛇菰的根茎会逐渐长大,伴随着寄主根也逐渐变粗。非常独特的是,根茎中的维管系统既包含蛇菰自身的,也包含寄主根的,二者形成复合的维管组织。这种独特的寄生方式确保了蛇菰与寄主之间的紧密连接,有助于蛇菰更好的从寄主获得养分。
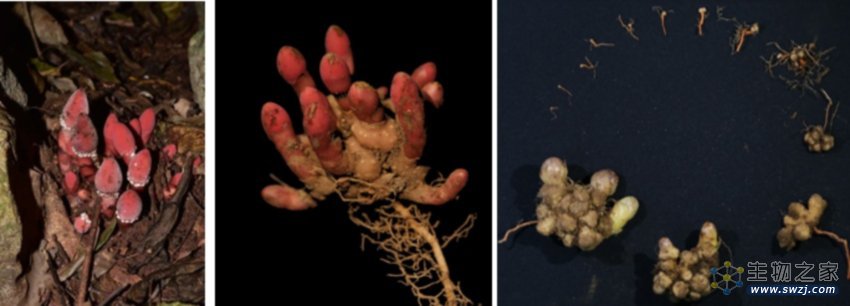
图1:蛇菰的形态及生活环境。左侧图:杯茎蛇菰的野外生活环境(拍摄者:魏泽,中国植物图像库),中间图:杯茎蛇菰的根茎、花茎、及寄主根的连接,右侧展示了球穗蛇菰不同的生长发育阶段
理解此类寄生植物的演化历程及生活机制,对我们探究物种起源、万物生长有重要意义。近日,深圳华大生命科学研究院联合昆明植物所、英属哥伦比亚大学等单位合作在Nature Plants发表了题为Balanophora genomes display massively convergent evolution with other extreme holoparasites and provide novel insights into parasite–host interactions的研究论文,完成了两种蛇菰的基因组解析,揭示了其独特形态和神奇生活方式的背后机制。

图2:发表文章截图
与自养植物相比,不同类群的寄生植物或多或少发生了基因的丢失。研究者首先找出了在多数自养植物中都保留的基因,然后分别判断各种寄生植物所丢失的基因比例。已经发表的文章显示,大花草科植物寄生花(全寄生)丢失了约44%的基因、旋花科植物菟丝子(介于全寄生和半寄生之间)丢失了约11.7%的基因、列当科植物松蒿、马先蒿和独脚金(半寄生)丢失了约2.4%-5.7%的基因,而全寄生的列当和埃及列当,分别丢失了13.11%-13.85%的基因。
那么,其它全寄生类群是否也发生了剧烈的基因丢失,不同类群的寄生植物是否发生类似的基因丢失,基因丢失与其环境适应之间是什么关系?这些都是植物科学领域非常关心的问题。
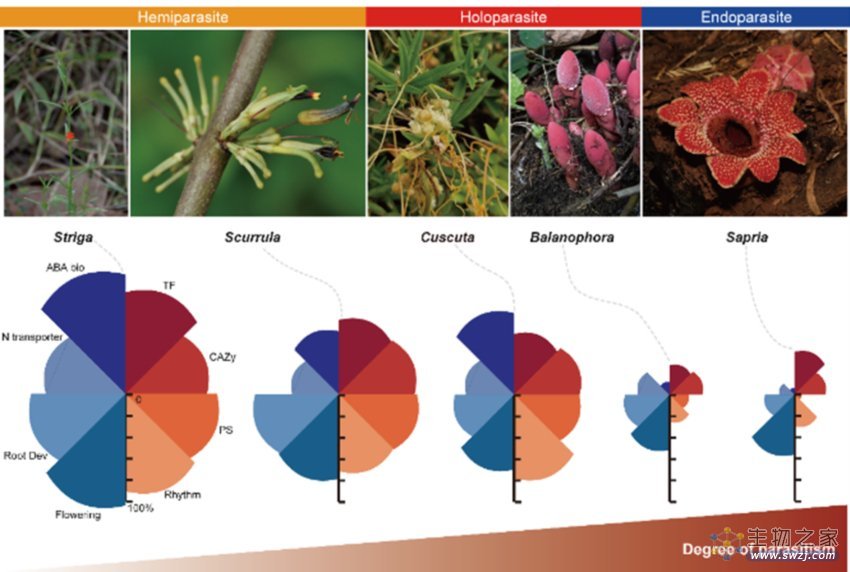
图3 来自5个类群的寄生植物的形态及一些关键功能的基因丢失
研究者对檀香目蛇菰科蛇菰属的两个物种杯茎蛇菰和球穗蛇菰进行基因组组装,组装大小分别为0.56Gb和1.15Gb,分别注释出14224和14283个蛋白编码基因。另外还组装了来自桑寄生科的半寄生植物小红花寄生,组装大小0.51Gb,共注释出38029个蛋白编码基因。
在被子植物类群中鉴定出保守的基因家族10277个,并进一步鉴定各寄生植物类群中发生丢失的基因家族,发现两个独立发生的寄生类群——蛇菰属及寄生花都发生了非常剧烈的基因丢失,分别有2898、2918、3925个基因家族丢失。而两个类群共同发生的基因家族高达2282个,分别占各自基因丢失的78.2% 和58.1%,表明在寄生程度高的全寄生植物,发生了趋同的功能的退化。而这些共同基因丢失,主要功能集中在光合作用和质体、色素代谢、昼夜节律、胁近响应、脱落酸的生物合成等功能,表明全寄生植物生活方式的转变可能极大的影响了这些功能。
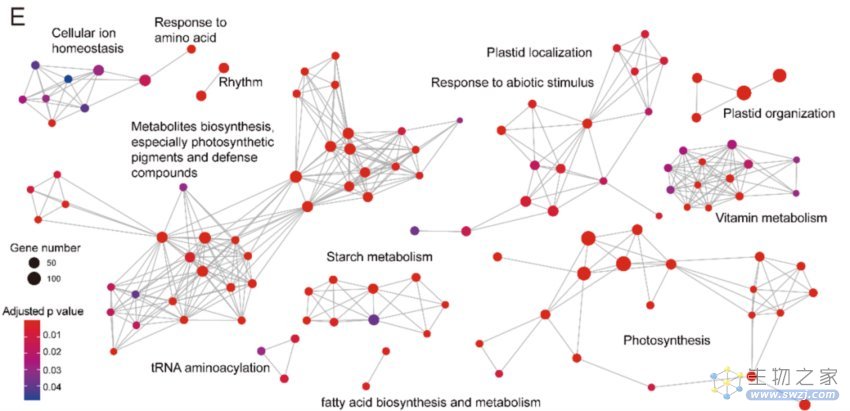
图4:蛇菰和寄生花中共同丢失基因的功能
除基因丢失外,蛇菰和寄生花还表现出多基因家族的强烈收缩。在被子植物中,一些基因家族大多表现为保留多个成员,且主要集中在信号传导、运输、以及代谢等功能上。通过对14个自养植物中保守的多基因家族进行分析,发现在蛇菰中,20.6%的基因家族完全丢失,50.0%的家族转变为单基因家族,仅有29.4%的基因家族仍然只留大于1个成员。寄生花表现出类似的变化。其中,蛇菰和寄生花仅保留483-634个转录因子(拟南芥:1720,葡萄:1544),甚至少于藻类植物的转录因子数目。
被子植物中的多基因家族,在蛇菰及寄生花中约有一半变成了单基因家族,表明基因成员增多所带来的有益影响,在寄生植物中减弱或完全消失。进一步对这些收缩的基因家族进行功能分析,发现功能主要集中在信号传导、对刺激的响应、器官发育、初生及次生代谢上。因而研究者推断,全寄生植物中高度退化的营养器官,代谢的减弱或丢失,以及寄主提供的相对稳态的环境,可能导致对生长及环境适应中复杂的调节及代谢系统的选择压力的减弱,从而导致基因家族的收缩。
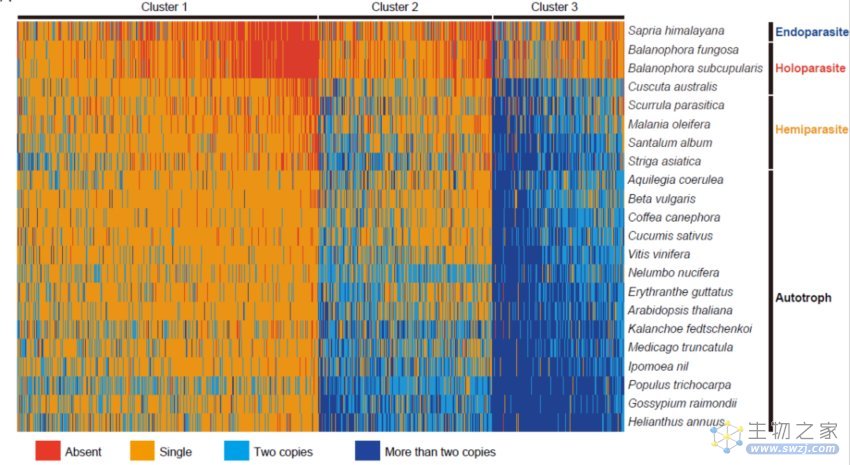
图5:根据基因基因家族中的基因数目,可以将自养植物中的基因家族分为三类,其中第1类在自养植物中多为单基因家族,第3类多为多基因家族,第2类处于中间状态。寄生植物在这三类中表现出不同的基因丢失水平
这些全寄生植物中保留下来的基因又发生了哪些变化呢?与自养植物相比,全寄生植物中保留下来的基因发生了非常高的同义突变和非同义突变。研究者检查了全寄生植物中的DNA修复基因,发现有一些DNA修复基因丢失,其总数也大大减少。
研究进一步推断,在寄生生活方式下,寄生植物的基因所受到的选择压力也将发生变化。通过对2729个保守的基因家族进行选择压力的分析,发现与自养植物相比,分别有51.9%和57.3%的基因在半寄生和全寄生植物中经历宽松的选择压力,而仅有9.5%和4.1%的基因在半寄生和全寄生植物中受到增强的选择压力。对各分枝及节点进行选择压力的分析,发现在大多数的寄生植物类群中,基因受到较宽松的选择压力。宽松的选择压力可能在多数寄生植物的基因进化过程中发挥主要作用,能够解释寄生植物的高的碱基突变速率、基因丢失、以及多基因家族的收缩。
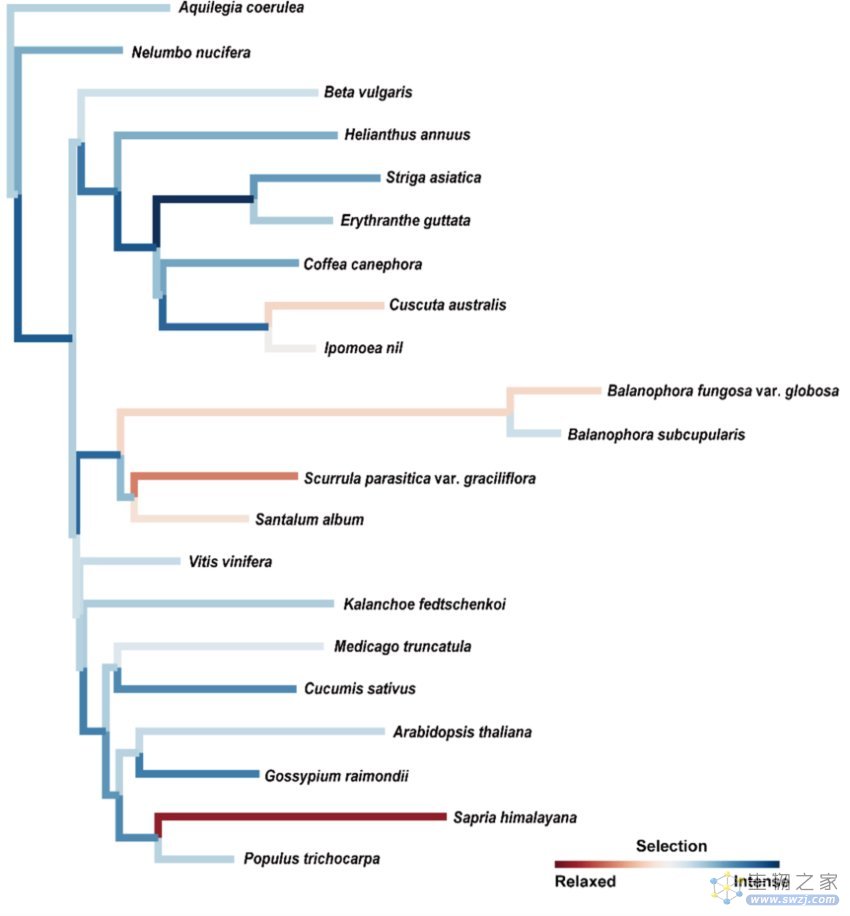
图6:各寄生植物及其近缘物种的基因所受到的选择压力的平均值。其中寄生植物类群大多表现出选择压力的放松
利用转录组数据分析了蛇菰与寄主之间的互作,包括mRNA在两者之间的穿梭,以及在蛇菰与寄主交接处发生的丰富的激素活动。例如在蛇菰可能在根茎中高表达了多个与水杨酸含量降低及信号抑制相关的基因。水杨酸作为一种植物激素,主要在免疫过程中发挥作用。病原体能够通过抑制水杨酸来削弱寄主的免疫反应,研究者推断蛇菰可能利用了相似的机制来降低寄主的抵御。
蛇菰属植物和寄生花都发生了脱落酸(ABA)关键合成通路的基因丢失,包括ABA1,ABA4,NCED,AAO3,其中AAO3基因编码ABA合成的最后一个酶,即使蛇菰可以利用寄主的中间体,当AAO3缺失时,也无法独立完成ABA的合成。而ABA信号通路的基因仍然被保留下来。研究者进一步研究ABA能否在蛇菰中被检测到,因而在蛇菰的花序、根茎、及寄主根中分别进行了ABA的含量测定,发现在蛇菰的花序中ABA的含量较高。可能蛇菰利用了其它未知的ABA合成通路,也有可能寄主中合成的ABA通过维管系统运输到蛇菰的花序中,并引发了蛇菰对ABA的响应。
全寄生植物在丢失光合作用能力后,大量的基因逐渐丢失,这一比例甚至高达38%,大大拓展了人们对于生物必需基因的认知。基因丢失作为进化的重要动力之一,一般被认为是中性的。但是某些基因的丢失可能对寄生植物适应其生活方式是有利的,例如寄生植物中开花调节基因的丢失、以及ABA合成基因的丢失,能够让寄生植物与寄主保持生理状态上的一致,更有利于其生存。寄生植物所受到的宽松的选择压力可能对其基因的进化发挥了主导作用,引起了基因突变的积累,以及基因的丢失。
深圳华大生命科学研究院陈晓丽、方东明,中国科学院昆明植物所许宇星为该论文共同第一作者。深圳华大生命科学研究院刘欢研究员,加拿大英属哥伦比亚大学Sean W. Graham 为论文共同通讯作者。该项目得到深圳市科技创新委员会基金的支持,获得深圳国家基因库对本项目数据产出和数据归档的支持。该项目是万种植物基因组计划的一部分(https://db.cngb.org/10kp/)。
全文链接:
https://www.nature.com/articles/s41477-023-01517-7