从生物学上来讲,三体人真的可行吗?
当然是可行的,我们不妨从生物演化的角度,详细分析三体人的进化过程。
地球上的生物,具有脱水耐性(Desiccation tolerance)的其实比较常见。无论细菌、真菌,还是植物和动物中,都有生物具有这样的能力。

我们重点来看看地球上具有此类能力的动物,与三体人的共性。
动物中的轮虫纲、线虫动物门、缓步动物门、鳃足纲,以及一些昆虫,都有物种存在很强的脱水耐性[1][2]。各类中的代表生物有:
轮虫纲——蛭形轮虫,从数万年永冻土中分离[3][4],能盗取基因[5],约8~9%外源基因。
线虫动物门——秀丽隐杆线虫,耐久型幼虫阶段可脱水98%[6],可在极端干燥环境存活4个月。
缓步动物门——水熊虫[7],可脱水99%,抗极端高低温、辐射,存活数年,有盗取基因能力。
鳃足纲——丰年虾[8],俗称卤虫,耐干旱高盐,幼虫脱水98%,最高可耐100%盐浓度。
昆虫——范氏多足摇蚊[9][10],又称昏睡摇蚊,脱水97%,长时间耐高低温,耐纯酒精。
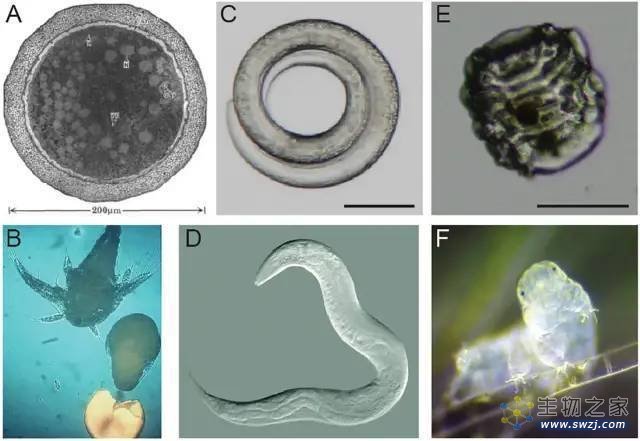
丰年虾、秀丽隐线虫、水熊虫两种状态对比(上下),图源:参考资料
当遇到极端环境,动物转变生理特征,转变为具有忍耐恶劣环境的状态,称为隐生(Cryptobiotic)。
在无脊椎动物中,隐生是很常见的现象,除了低湿隐生(脱水耐性)外,还有低温隐生、高温隐身、缺氧隐生、变渗隐生等等。
不难发现,具有脱水耐性的生物,往往具有多种隐生能力。
在《三体》原著中,三体人其实也不仅仅具有脱水耐干旱的特性,也同时能耐高温、低温、耐辐射。三体人脱水后,变成干纤维,除了可以等待复活外,也可以一把火烧掉,成为燃料。三体人判处死刑,或者年老、找不到工作,就是脱水一把火烧掉。
这不仅说明了三体人的碳基属性,也说明了它们脱水后,具有极低的含水量。
三体人与地球生物的高共性,适宜地球环境,也是他们意图攻占地球的根本原因。
通过分析以上生物的共通生理特性,我们能够探索出三体人的进化秘密。具有极端环境抗性的地球生物,它们身体内大多具有相同的秘密——海藻糖(trehalose)[11]。
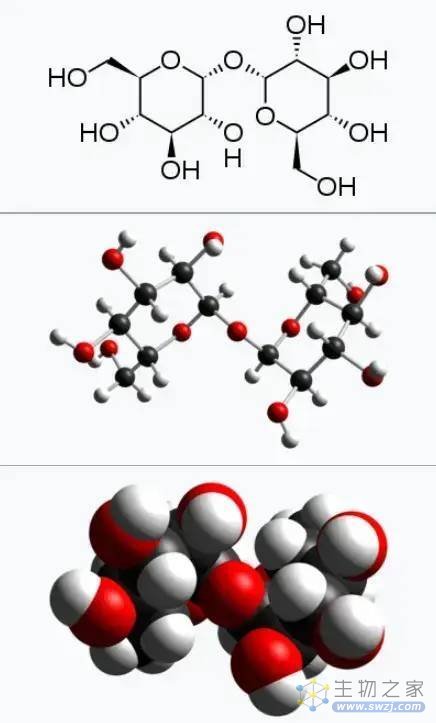
虽然细胞中的一些蛋白质、多不饱和脂肪酸、多胺,以及其它的一些糖分,也有利于脱水耐性[12][13],但海藻糖是这些生物,具有极端环境抗性的主要共同物质[14][15],不仅仅是动物,就连具有脱水耐性的真菌(如香菇)和植物(如卷柏),也主要是海藻糖在发挥作用。
海藻糖具有什么样的神奇魔力,才让这些地球生物,共同选择了这种物质?
因为海藻糖在干燥环境下,会发生玻璃化,形成非晶态物质。
海藻糖玻璃化状态下,没有尖锐晶体,不仅可以避免对细胞结构的破坏,还能降低胞内水蒸汽压力,增加对脱水的抵抗力。同时玻璃化的粘稠状态,也能减少分子扩散,基本停止细胞内的所有生化反应。同时还会抑制脂质融合,对细胞膜和细胞内蛋白质都具有稳定和保护作用,从而确机体保完全休眠和长时间的稳定[16][17]。
脊椎动物脱水耐性低,正是缺少这样的机制。
在脊椎动物中脱水耐性最强的是红斑蟾蜍,最高也只能达到40%。低温耐性最强的是林蛙,主要利用体内的尿素和葡萄糖作为低温保护剂,可在零下几度身体三分之二以结冰状态存活[18]。
但这些特征比起隐生的无脊椎动物都差了很多。
更高等的脊椎动物,无论血液循环系统、免疫系统、神经系统,各类器官都高度依赖水环境,往往没有脱水耐性。海藻糖在人体中不仅比例低,而且主要促进细胞自噬[19],并不发挥对极端环境耐性的作用。
这也注定了三体人与地球人截然不同的生理构造。
无脊椎动物之所以广泛用海藻糖作为隐生状态的保护剂,并不是它们专门应对隐生而进化来的,而是它们的“血糖”正好就是海藻糖。
即便不能隐生的昆虫,绝大多数都能忍耐干旱,轻轻松松可脱水50%存活,超过蛙类的极限,而且体型更大的昆虫,表现出更高的脱水耐性[20]。
所以从地球生物推测,三体人的血糖,大概率就是海藻糖,而且它们的血糖浓度可能高于地球人。根据地球生物的特征,可以猜测,三体人在脱水的过程中,体内超过20%以上的干重物质都会转化成海藻糖。
作为大型生物,仅仅只有海藻糖的血糖,还不足以具有极高的脱水耐性。
三体人还应该具有什么样的特征?
通过分析上面的多种地球生物,不难发现,较高等的无脊椎动物中,鳃足纲和昆虫纲属于节肢动物,它们和缓步动物门都属于泛节肢动物。
泛节肢动物普遍存在几丁质(甲壳素,属于一种多糖)外骨骼,同时也在地球生物史中出现过大型化的生物。
除了脱水耐性,三体人在炙热环境下还需要反射太阳辐射,那么三体人是必然存在外骨骼的。
不过外骨骼的进化路线是多种多样的,仅仅钙化路线就多达18种,并不一定需要考虑节肢动物的外骨骼[21]。

钙的运用可以增加外骨骼的强度[22],几丁质的运用可以增加韧性和弹性[23],二氧化硅的运用可以增加对太阳辐射的反射[24]。当然,不排除可以像鳞足类那样运用黄铁矿,具有含铁的外骨骼[25]。
这样三体人的外骨骼大约具有三层,最外层为二氧化硅,这一层可能很薄,类似于昆虫外骨骼的蜡质分子。中间是矿化层,可能含钙和铁等等,具有很高的强度。应该存在微孔,可以运输硅化物到外骨骼表面。内部是几丁质,附着肌肉,不仅保护内部脆弱的组织和器官,还能给肌肉储存弹性势能。
三体人可能存在的相似肌肉系统
除了外骨骼外,还可以通过呼吸和排泄来降低水分的流失。
三体人是怎么呼吸的?
考虑到有氧呼吸的种种优势,三体人应该也属有氧生物。
至于它们的呼吸方式,需要从以下两点来看:
1. 通过三体人的思维来看,中枢神经系统应该也是集中的,而不是像章鱼神经系统那样分散。
2. 三体人的神经系统可以一开始就是集中的,就像脊索动物门一样;也可以是分散,在不断进化过程中,逐渐变得集中,例如早期可以类似于环节动物、昆虫那样的链状神经系统。
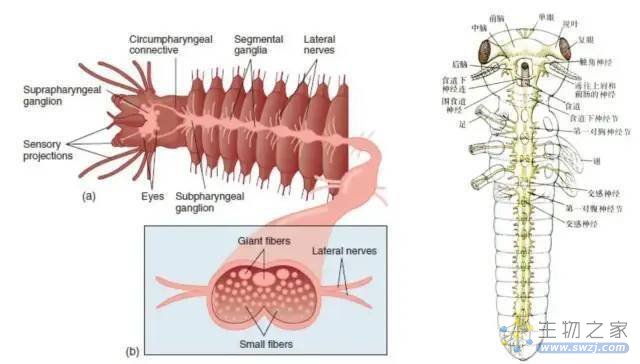
考虑到三体人生活的恶劣环境,当它们在进化史上还属于低等生物时,用肺这样的器官呼吸是具有很大代价成本的。很有可能,大多低等的三体人(或者三体文明中的低等动物)全身分布着类似气门这样的呼吸器官。

气门上具有覆盖物,可以减少水分的流失。
在原著中,三体人经过近200次文明,它们的进化次数可能比这还多得多。
那么在反复往高等生物进化的过程中,随着神经系统、循环系统、骨骼系统变得越来越发达,它们可能就像陆地脊椎动物退化掉鳃一样,退化气门,演化出具有高效呼吸的“肺”。当然,它们肺的具体形式和人类会有差异,同时具有抗旱、冷、热等特性。
由于三体人还需要进入隐生状态,那么三体人的气门可能并没有完全退化。
当环境变得恶劣后,它们转化体内有机质为海藻糖的同时,还会关闭肺部,同时保持气门通畅,持续极其微弱的呼吸状态。
通过极低的新陈代谢,保持机体的生命状态。
三体人的繁殖和交流方式,又可能存在怎样的进化机制?
按照原著,三体人的繁殖方式是父母的有机质互相融合,然后诞生3~5个后代,后代会继承父母的部分记忆。
那么,三体人的这种生殖方式,相当于低等生物的接合生殖+分裂生殖。
地球上高等生物的两性异配生殖方式,其实是高度依赖外部水环境所进化而成的。三体人可能经常面临极端缺水环境,那么主要的繁殖方式沿着融合-分裂路线,是合理的。
就像地球生物的异配生殖可以逐渐进化成哺乳动物这样的复杂生殖方式一样,三体人融合分裂的生殖方式,也可以进化到高等的形式,可能在生殖过程中,机体也存在复杂的生化变化,组织和器官有着自身的融合和分裂规则,这里就不详细探讨了。
三体人如何才能做到继承父母的部分记忆?
这一点上,我们可以参考昆虫所具有的成虫盘。
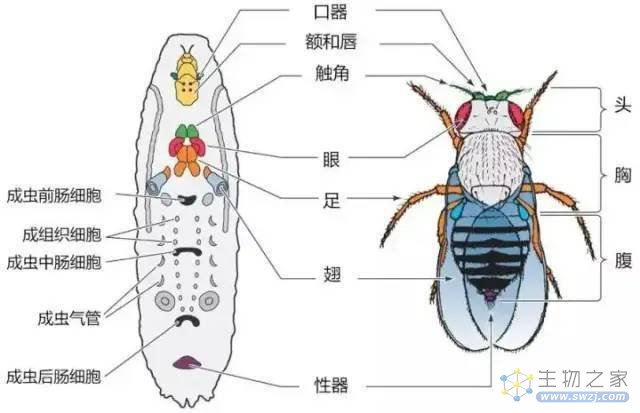
完全变态的昆虫,幼虫体内具有典型的成虫盘。
幼虫只管不断地吃,化蛹之后,成虫盘内的干细胞就会再分化成成虫的各个器官。
所以才有人认为,成虫是寄生在幼虫体内的,最终取代幼虫。幼虫不断吃吃吃,只是给成虫做了嫁衣。但这种说法,其实并不严谨。
按照成虫盘形成的机制,不难猜测,三体人之所以能继承父母的部分记忆,具有相似的过程。
在繁殖过程中,父母承载记忆的神经系统并不会直接被破坏,而是经过融合分裂后(不排除有复制的可能),被后代所继承,会类似于成虫盘一样被保护起来。
在幼体不断成长的过程中,随着神经系统的发育,功能逐渐完善后,这些记忆就会不断的解开,并和后天记忆一样,成为自己记忆的一部分。
至于三体人单纯运用脑电波进行交流,这对进化的挑战,具有最高的难度。
这种不依赖于五官的交流,有些类似于昆虫的信息素。
当然直接以脑电波交流,的确才是最底层的机制。
因为无论听觉、触觉、视觉等等各种感官,本质上的力都是电磁力。而且外部信号的转化,本质上也都是转化成电信号传导的。
我们可以推测,三体人神经系统的底层逻辑和地球生物是相似的。但他们神经系统输出和接受信号的微观结构,会存在调制解码系统。
虽然进化出来的确难上加难,但原著设定中三体人进化出文明就有200多次。
他们的生物演化史,比地球生物还要内卷。
不排除在最后一次进化过程中,恰好出现了特定的环境和相应的生理构造,进化出脑电波交流的可能。虽然概率很低,但也并非没有可能。
有了这样的神经系统,三体人的社会形式就很好解释了。
地球上,单纯因为信息素交流,蚂蚁之类的昆虫就可以形成庞大而紧密的社会结构。纯信息素的控制,也让蚂蚁没有“情感”。
当然蚂蚁没有情感,也可能是神经系统过于原始的体现。而且三体人对个体,也仅仅是交流,并不会像蚂蚁那样通过信息素控制个体。

这也是为什么,三体人并不会形成昆虫那样的虫族社会体系。
虽然三体原著提到三体社会是绝对理性的,但其实三体人个体是具备各种情感的。例如原著中的三体人个体就有表现出一些情绪、情感反应。
我们可以这样理解,三体人是具备情感,但是被高度压抑的。
人体社会被要求绝对理性,这是他们应对极端环境,进化出来的生存策略。如果他们真的能生存在地球上或者与地球人长期接触,这样的绝对理性是可能被瓦解的。
三体人社会属性上的理性,其实也是体现在透明交流带来的高压。可以想象,从现在开始地球人的所有思想都变得完全透明,那么任何人的表达都会变得谨慎,情绪和情感自然就受到了压抑。
个性被限制后,一个集体更容易形成共同的声音。
任何高效率的组织,都必须有领袖。这种共同声音,就会集中在领袖上,形成权力的集中。再加上,三体人通过统一行动渡过一次次的灾难,随着领袖威望的增加,形成专制元首制的可能性就会越来越大。
以上便是三体人可能的进化过程。
我们不难发现,虽然三体人具有多种地球生物的特征,但并不能在地球上找到十分相似的生物。
不过,按照原著的设定,三体人的生理特征,其实是比较符合进化逻辑的。
不得不说,在三体人的设计上,大刘是很用心的。
参考文献:
[1] Hibshman J D, Clegg J S, Goldstein B. Mechanisms of desiccation tolerance: themes and variations in brine shrimp, roundworms, and tardigrades[J]. Frontiers in Physiology, 2020, 11: 592016.
[2] Watanabe M. Anhydrobiosis in invertebrates[J]. Applied entomology and zoology, 2006, 41(1): 15-31.
[3] Shmakova L, Malavin S, Iakovenko N, et al. A living bdelloid rotifer from 24,000-year-old Arctic permafrost[J]. Current Biology, 2021, 31(11): R712-R713.
[4] Ricci C. Anhydrobiotic capabilities of bdelloid rotifers[J]. Hydrobiologia, 1998, 387: 321-326.
[5] Gladyshev E A, Meselson M, Arkhipova I R. Massive horizontal gene transfer in bdelloid rotifers[J]. science, 2008, 320(5880): 1210-1213.
[6] Erkut C, Vasilj A, Boland S, et al. Molecular strategies of the Caenorhabditis elegans dauer larva to survive extreme desiccation[J]. PloS one, 2013, 8(12): e82473.
[7] Møbjerg N, Halberg K A, Jørgensen A, et al. Survival in extreme environments–on the current knowledge of adaptations in tardigrades[J]. Acta physiologica, 2011, 202(3): 409-420.
[8] Gajardo G M, Beardmore J A. The brine shrimp Artemia: adapted to critical life conditions[J]. Frontiers in physiology, 2012: 185.
[9] Sogame Y, Kikawada T. Current findings on the molecular mechanisms underlying anhydrobiosis in Polypedilum vanderplanki[J]. Current opinion in insect science, 2017, 19: 16-21.
[10] Hinton H E. Cryptobiosis in the larva of Polypedilum vanderplanki Hint.(Chironomidae)[J]. Journal of Insect Physiology, 1960, 5(3-4): 286-300.
[11] Ramløv H, Westh P. Survival of the cryptobiotic eutardigrade Adorybiotus coronifer during cooling to− 196 C: effect of cooling rate, trehalose level, and short-term acclimation[J]. Cryobiology, 1992, 29(1): 125-130.
[12] Tunnacliffe A, Lapinski J, McGee B. A putative LEA protein, but no trehalose, is present in anhydrobiotic bdelloid rotifers[J]. Hydrobiologia, 2005, 546(1): 315-321.
[13] Shen‐Miller J, Mudgett M B, Schopf J W, et al. Exceptional seed longevity and robust growth: ancient sacred lotus from China[J]. American Journal of Botany, 1995, 82(11): 1367-1380.
[14] Tunnacliffe A, Lapinski J, McGee B. A putative LEA protein, but no trehalose, is present in anhydrobiotic bdelloid rotifers[J]. Hydrobiologia, 2005, 546(1): 315-321.
[15] Erkut C, Vasilj A, Boland S, et al. Molecular strategies of the Caenorhabditis elegans dauer larva to survive extreme desiccation[J]. PloS one, 2013, 8(12): e82473.
[16] Green J L, Angell C A. Phase relations and vitrification in saccharide-water solutions and the trehalose anomaly[J]. The Journal of Physical Chemistry, 1989, 93(8): 2880-2882.
[17] Elbein A D. The metabolism of α, α-trehalose[J]. Advances in carbohydrate chemistry and biochemistry, 1974, 30: 227-256.
[18] Storey K B, Storey J M. Frozen and alive[J]. Scientific American, 1990, 263(6): 92-97.
[19] Aguib Y, Heiseke A, Gilch S, et al. Autophagy induction by trehalose counter-acts cellular prion-infection[J]. Autophagy, 2009, 5(3): 361-369.
[20] Renault D, Coray Y. Water loss of male and female Alphitobius diaperinus (Coleoptera: Tenebrionidae) maintained under dry conditions[J]. European Journal of Entomology, 2004, 101(3): 491-494.
[21] Porter S M. Seawater chemistry and early carbonate biomineralization[J]. Science, 2007, 316(5829): 1302-1302.
[22] Nedin C. Anomalocaris predation on nonmineralized and mineralized trilobites[J]. Geology, 1999, 27(11): 987-990.
[23] Bennet-Clark H C. The energetics of the jump of the locust Schistocerca gregaria[J]. Journal of Experimental Biology, 1975, 63(1): 53-83.
[24] Falasco E, Bona F, Badino G, et al. Diatom teratological forms and environmental alterations: a review[J]. Hydrobiologia, 2009, 623(1): 1-35.
[25] Sun J, Chen C, Miyamoto N, et al. The Scaly-foot Snail genome and implications for the origins of biomineralised armour[J]. Nature communications, 2020, 11(1): 1-12.