01
昂贵的性
当我们谈到性时,你会想到什么?美好与爱情?还是风险和威胁?无论情感将你带往何方,我们都得认同的一个直截了当的简单事实——性是我们生育后代的必需途径。
但我们为什么需要通过性来制造婴儿,而不能简单地进行自我克隆呢?事实上,在性这方面,人类一点都不特别——地球上的大多数动物都是有性生殖物种。一项调查发现,只有22种鱼类、23种两栖动物和29种爬行动物的生殖无需通过性——这在已知的42,000多种脊椎动物物种中,只占取了非常小的一部分 。
性是非常昂贵的。这里指的并不是一个男人或女人为了赢得梦中情人的爱可能要耗费的时间、金钱或心机策略——这些是社会科学家和言情小说家所需要考虑的。在演化生物学家眼中,性的代价更为现实。
效率低
首先,与无性生殖相比,有性生殖效率极低——事实上,有性生殖的效率只有无性生殖的一半。试想一下这样一个场景(见下图):假设无论是通过有性生殖还是克隆生殖,一个雌性动物每年能生育两个后代。
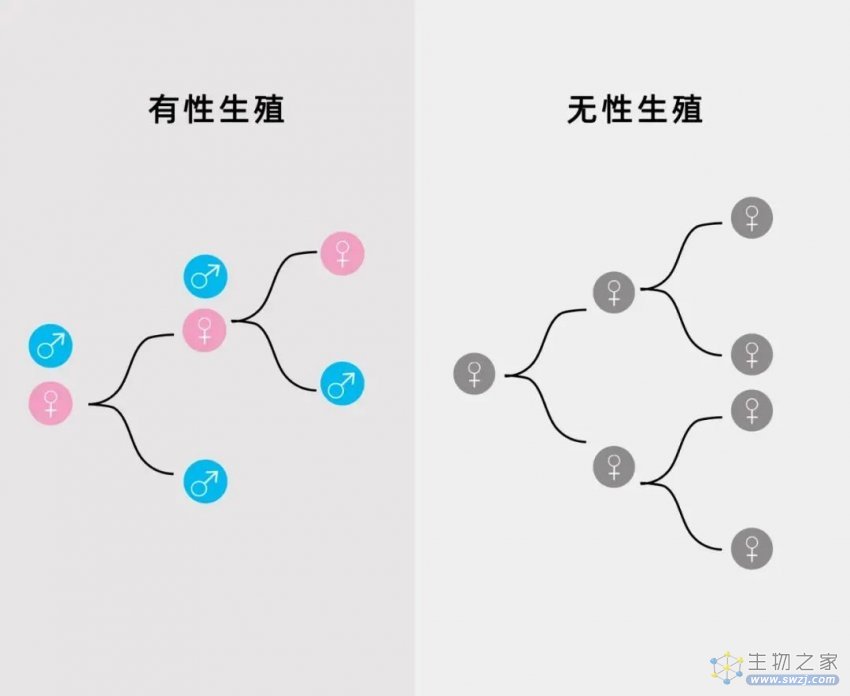
在有性生殖的情况下,该雌性动物(红色)会分别育有一个雌性(女儿)和雄性(儿子)后代,这意味着它只有一半的后代(女儿)可以直接促进种群数量的增长;但在克隆生殖的情况下,无性生殖的雌性(灰色)能复制出自身的两个副本,因而它们两个都可以通过自我复制继续制造出雌性孙辈。在这种情况下,有性生殖的种群数量保持不变,而克隆生殖的种群数量会指数增长。
速度慢
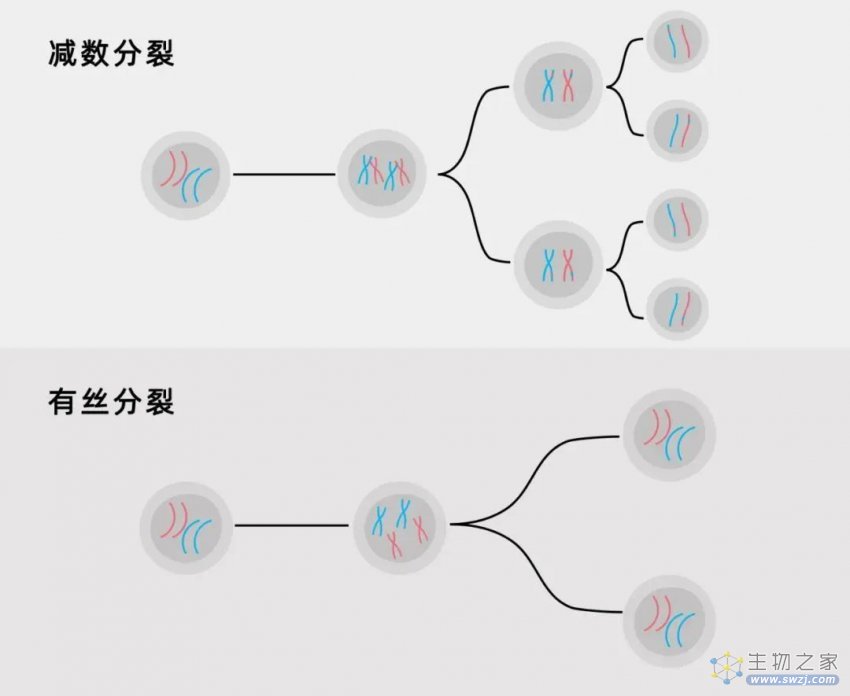
除此之外,有性生殖的速度也慢于克隆。在有性生殖中,细胞必须经历一个名为减数分裂的复杂过程。这包括染色体的交叉和遗传物质的重组,通常需要超过十个小时的时间。相比之下,克隆生殖所需要的细胞过程则更为简单,这一过程被称为有丝分裂,只需要直接复制基因组和一个细胞分裂成两个完全相同的子细胞,它可以在15分钟之内完成。对于那些需要很长时间才能达到成熟期,且在两个连续的生殖事件之间需要长时间间隔的物种(例如人类和其他大型多细胞动植物)来说,时间成本是微不足道的;但是对于微观的生物,尤其是单细胞生物来说,这种时间成本是巨大的。
冲突多
性的最显著代价是通过生态相互作用、性选择、配偶竞争和性冲突体现的。雄孔雀华丽的尾屏对雌孔雀极具吸引力,但伴随而来的代价是增加了死亡风险,因为美丽的颜色不仅能吸引配偶,也能吸引捕食者,而且尾屏的重量和尺寸也使雄孔雀在逃离危险时更加困难。此外,许多种类的雌性蜘蛛和螳螂在交配前、交配期间或交配后会吃掉它们的配偶。更令人惊讶的是,雄性的宽足袋鼩——一种小型的与鼩鼱类似的有袋类动物——会毫不夸张地交配致死。在长达2~3周的交配期开始之前,雄性的宽足袋鼩就会停止制造精子,准备全力投入马拉松式的交配竞赛。在此期间,它们会疯狂地寻找雌性并试图与尽可能多的雌性交配。据观察,它们会连续几个晚上不间断地交配6~12个小时,很少或者根本没有时间寻找食物。随着交配期的进行,雄性宽足袋鼩的体重随之减轻、皮毛脱落、双目失明、出现内出血,最后因免疫系统完全崩溃而痛苦地死去,而这一切都发生在它们一岁生日的几周前。这听起来或许疯狂,但这种“自杀式繁殖”恰恰是自然选择为最激烈无情的交配竞争量身定做的解决方案。
有性生殖既需要雄性也需要雌性,由于两性对生殖成功的需求不同,因此经常出现两性冲突,从而加重有性生殖的成本。
对雄性来说,最优策略通常是与尽可能多的雌性交配,因为制造精子是个相对廉价的过程。由于雄性动物——尤其是哺乳动物很少帮助雌性分担抚育后代的工作,它们竞争的实际上是由当地所有雌性动物所生养的有限数量的后代的父权。对于雌性来说,由于产卵和照顾后代的成本要高得多,因此它们更感兴趣的是选择一个基因质量高或资源丰富的配偶(例如能保护觅食领地和/或保护雌性免受其他雄性的骚扰)。
事实上,由于交配会耗费本可用来寻找食物或照顾后代的时间,而且可能会吸引捕食者并增加感染性病的风险,因此雌性通常会主动避免或抵抗雄性的交配企图。在极端情况下,两性冲突会导致雄性对雌性造成直接的身体伤害,或者通过杀婴来对雌性造成间接伤害,从而迫使雌性停止对其他雄性后代的付出,并更快地恢复繁殖能力。
杀婴听起来很残忍,但对某些物种的雄性来说却是一种有效获取父权的方式,甚至有可能是唯一的有效方式。
我们可以以狮子为例来说明这一点。狮子的妊娠期需要将近4个月的时间,母狮会在幼崽18个月大时才会恢复性行为。但是一旦母狮失去了她的幼崽,她可以在几天到几周内就恢复交配和受孕能力。相比于母狮将幼崽抚育到独立最少要花费的22个月的时间,一组雄狮(联盟)能够控制自己领地的时间是非常短的。对于只有1~2只雄性的小联盟,统治期通常不到25个月,即使是有4~6只雄性的大狮群,统治期也很少超过50个月。因此对狮子来说,留下自己的亲生骨肉,是一场与时间赛跑的比赛,只有那些设法让母狮迅速恢复繁殖能力(通过杀死依赖它们的幼崽)的雄狮才能在演化的战场上取得成功。
02
性别的演化
在考虑到有性生殖与自我克隆相比所具有的(通常是巨大的)成本之后,如果你开始困惑于为何性在自然界中如此普遍,那么恭喜你,你已经在像演化生物学家一样思考了。
事实上,性别的演化是一个长期存在的谜题,它隐藏了太多的谜团,以至于被称为“演化生物学问题中的女王”。如今,关于地球上的所有真核生物的共同祖先(大约可追溯到十亿年前)都是有性生殖的这一点,已得到了人们的普遍共识,因为相同的基因和涉及到减数分裂(有性生殖中最重要的一步)的基本细胞机制,都出现在了生命之树的所有主要真核生物的分支中,包括动物、植物、真菌。
尽管代价巨大,但由于有性生殖已经长期存在而且广泛分布在生命树中,它所带来的利益一定更大。然而,这些益处的具体形式仍有待商榷。迄今为止,最具影响力的理论有两个,要么与有害突变在无性物种中的积累有关,要么与有性物种在宿主-寄生的共同演化中的优势有关,它们分别被简称为“穆勒棘轮”(以诺贝尔奖得主赫尔曼·约瑟夫·穆勒命名)和“红皇后假说”。这两种理论都是基于有性物种和无性物种之间的一个基本区别:无性生殖的后代都是彼此相同的,而有性生殖的后代在性状和基因组上都表现出更大的多样性。
穆勒棘轮
“穆勒棘轮”理论意识到,尽管DNA的复制大体上十分准确,但它仍有可能因基因突变而出现错误,这些突变大多都有害。通过克隆生殖的物种所产生的子代,虽然与亲代几乎有着相同的基因,但仍可能出现少量错误。这些新的错误将被保留在下一代的子代繁殖过程中,循环往复、代代相传,就像一个棘轮的运动一样——单向且不可逆转。经过一代又一代之后,这些小的错误越积越多,导致的结果就是后代比祖代要弱得多。
但是对有性生殖的物种来说,有害突变的积累是可以消除的。当然,在卵子和精子的制造过程中,仍会有突变发生,但是这些突变的发生是随机的,而且往往位于基因组的不同位置。因此,当一个卵子与精子结合以产生后代时,遗传物质的重组会产生一些拥有很多有害突变的不幸后代,以及一些没有突变的幸运后代。有时,甚至有些抽中上上签的后代还会获得一些有益突变。在自然选择下,遗传质量最好的后代有更高的生存和繁殖机会,而那些具有大量有害突变的后代将被剔除出基因库。这样一来,有性生殖的物种就能维持甚至提高它们的遗传质量,因此从长远来看,相比于会遭受突变积累和基因组退化的棘轮效应的无性生殖物种,有性生殖物种更具竞争力。
红皇后假说
另一个最有影响力的理论是“红皇后假说”,它是以刘易斯·卡罗尔(Lewis Carroll)笔下的童话故事《爱丽丝镜中奇遇记》中的红皇后命名的。
在这个故事中,爱丽丝与红皇后手拉手奔跑,当她跑到上气不接下气时,发现周围的树和其他物体根本没有改变位置。惊讶且困惑的爱丽丝告诉红皇后,在她的家乡,如果她飞快地奔跑一段时间的话,就会抵达别的地方,而不是仍然待在原地。红皇后听了回答说:“现在,在这里(镜中之国),以你的速度你只能逗留在原地”。演化生物学家常用红皇后的竞赛来类比宿主与寄生虫之间的共同演化。
试想一下通过克隆生殖的物种。最初,种群会得到迅速增长,并且很快充满相差无几的个体,直到出现一种致命的寄生虫(如结核菌、艾滋病毒、疟疾等)。只要种群中的一个个体能被这种寄生虫能杀死,那么其余个体也在劫难逃,因为所有克隆个体的遗传弱点都是完全相同的。在这种情况下,寄生虫会像野火一般蔓延,直到消灭宿主物种的每一个成员。
但在有性生殖的物种中,同样的致命寄生虫却通常无法传播很远。因为所有的个体都不同,因此这种寄生虫迟早会遇上某些有天然抵抗力的个体。如果这些寄生虫还想要继续蔓延到其他种群,它们就必须历经时间来变异和演化。不过,在这段时间内,有性生殖已经让那些抗寄生虫的宿主对其种群的基因库进行更新重组,并产生了与自身完全不同的后代。这使得一些后代再次领先于致命寄生虫一步。这就好比在镜中的爱丽丝无论怎样全力奔跑,她和周围环境的相对位置也只能保持不变,宿主想要生存下来,并要始终比同样也在快速演化的寄生虫稍微领先一点一样,它们需要有性生殖来对自己的基因组进行不断地重组,从而创造出不同的后代。
两面下注
需要注意的是,这两种理论并不互相排斥,而是互补的。尽管“性别的演化”是一个存在已久的研究领域,但它仍在迅速发展,总有新的理论被不断地提出。例如2017年,有科学家提出了一个新的理论,认为有性生殖是一种风险分摊策略,称为“两面下注”。这一新理论既考虑了有性生殖的负面影响(后代数量少),也考虑了有性生殖的益处(后代多样化)。
我们将有性生殖和无性生殖之间的竞争比喻为一种购买彩票的场景。无性生殖就像买大量号码相同的彩票,而有性生殖就像是买少量的彩票,但每张彩票都有不同的号码。因为中奖号码是不可预测的,所以押注在同一组号码上有可能获得巨大的收益,但更大的可能是全盘皆输。相比之下,在不同的号码上押注永远无法带来太大的回报,但多数情况下也不会出现彻底的损失。同样,由于环境上的波动是不可预测的(谁知道下一个春天是温暖还是寒冷、湿润还是干燥?),一旦环境超出了无性生殖个体的耐受性范围,整个种群就都会灭绝。相比之下,有性生殖物种产生的后代虽然数量更少,却更加多样化。因此,无论环境如何变化,总有幸存者能够活到下一代。有性生殖的“两面下注”(或简单说就是不孤注一掷地将“所有鸡蛋都放在一个篮子里”)理论为有性生殖的演化提供了一层额外的解释。
03
性别本身
到目前为止,我们一直在讨论有性生殖的成本与收益。现在,让我们跳出成本与收益的账簿,将思绪回到这一过程本身。若要有性生殖发生,我们需要一个雄性和一个雌性配成一对,然后……但先等等!为什么需要一个雄性和一个雌性?到底什么是雄性和雌性?
雌与雄
事实上,这种看似“幼稚”的问题其实并不容易回答。雄性与雌性的定义与外表无关:对有些物种来说,这两种性别看起来几乎是完全相同的;而对于另一些物种,例如折衷鹦鹉,雄性和雌性的颜色虽然差异巨大但同样绚丽多彩。除了外表,这与体型也无关:对哺乳动物来说,雄性动物通常更大,但对鱼类、蜘蛛和许多昆虫来说,雌性可能比雄性大得多。除此之外,这与统治力和攻击性、与抚育后代的投入程度,甚至与性染色体都无关——许多物种根本就没有性染色体,它们的性是由发育过程中的温度和其他环境因素决定的,鳄鱼和海龟就是例子。
让人意外的是,雄性和雌性之间最重要的区别在于他(她)们产生的生殖细胞:雄性是产生小且可移动的生殖细胞(精子)的性别,而雌性是产生大而营养丰富的生殖细胞(卵细胞)的性别。但即使是这样一般化的定义,也只适用于只有两种不同性别的物种。(是的,有些物种具有两种以上的性别。迄今为止的冠军是一种蘑菇裂褶菌,它有23000多种不同的性别!)能同时产生精子和卵细胞的物种被称为雌雄同体。
灰色地带
“雌雄同体”一词的英文 hermaphrodites 是以希腊神话中的赫马佛洛狄忒斯(Hermaphroditus)和他/她略带悲剧色彩的故事命名的。
在希腊神话中,赫马佛洛狄忒斯在出生时是个小男孩,他快乐地长大,成了一名非常俊美的青年。直到有一天,一个水仙女爱上了他。水仙女对他的爱既深切又狂热,她祈祷能与他结合,永不分开,一位(行为上更像一个疯狂生物学家的)更强大的神回应了她的祈祷,真的将他们照字面意思所说的那样合二为一,所以自那之后,赫马佛洛狄忒斯(现在的被永远结合永不分开的夫妇)就拥有了男性的生殖器和女性的身体。
赫马佛洛狄忒斯的故事或许听起来荒谬,却也告诉我们一些与雌雄同体有关的真实情况:如果你是一个孤独生活的个体,很少有机会遇到一个潜在的伴侣(就像水仙女那样),一旦你遇到了同类的成员,那么最好不要在意性别,重点是能够交配。不管对方有的是精子还是卵细胞,都可以通过成为雌雄同体来确保生殖的可能,做好迎接两种性别的准备。
你可能会想,“水仙女很聪明嘛”,但生物学也为她添加了一个悲剧性的转折——雌雄同体通常不能自我受精,所以即使她与她的所爱看似永远地结合在了一起,他们却永远不可能真正地结合。
另一个显而易见的悲剧之处在于——一旦你变成了雌雄同体,那么你将一直处于灰色地带,无论对雄性还是雌性都不太可能显得有多大吸引力。就像样样皆通的人往往样样稀松,雌雄同体既不能很好的执行雄性功能也不能很好地执行雌性功能,除非在很少能遇到配偶的特定环境下,它们永远无法胜过有专属性别的个体(清楚的雌性与雄性)。
事实上,自然界中的雌雄同体(或确切地说是同时既是雌性又是雄性的个体)通常是行动迟缓且孤独生活的无脊椎动物,比如蜗牛、蛞蝓和蚯蚓。
性的转变
机敏的读者可能要问,如果雌雄同体难以出众,为什么不先以专门的一种性别存在,在需要时再换成另一种呢?
是的,许多物种确实演化出了这种策略,最常见的是在鱼类中,它们的性别变化方向通常与婚配制度有关。在一夫多妻制下,个体通常从雌性开始,然后当它们长大到能够管控一片领地和一整个雌性后宫时,就会变成雄性;而在一夫一妻或随机交配的系统里,个体通常以雄性开始,再向雌性转变,因为大的雌性的卵子在数量与质量上都比小的更优,但即使是小的雄性也能产生足够好的精子供其配偶受精。
尽管变性是一个耗时耗力的过程,但对于那些捕食风险非常高的物种来说变性是件高回报的事。
例如生活在珊瑚中的鰕虎鱼,如果两条性别相同的鱼发现它们身处同一个珊瑚岬,这时比起游到几米之外的另一个珊瑚岬去寻觅不同性别的个体,变性是种更为谨慎且在演化上更为成功的策略。对于鰕虎鱼这类小鱼来说,包围在珊瑚生态系统周围的是许多饥饿的大嘴,即使只是游上几米也能威胁到生命,因此变性成为了值得妥协的一件事。
然而最奇特的变性物种,是一种海洋中的多毛类蠕虫,叫 Ophryotrocha puerilis 。这种蠕虫是一夫一妻制的,最初,一对蠕虫中较大的那只会扮演雌性的角色,而小的那只成为雄性——这是有意义的,因为雌性越大越能产生更多更好的卵子,而雄性不需要多大就能产生足够的精子来为这些卵子授精。但是,由于制造卵细胞的成本远高于制造精子,因此雄虫会生长得更快,最终变得比雌虫更大。然后它们就会同时变性,大的那个从丈夫变成了妻子,接管了生产卵子的繁重工作,直到另一只再次变大。这一循环不断更替,直到死亡将这对平等而富于奉献的夫妻分开。
04
未知的神秘
虽然性别演化这一研究领域一直在不断地积极前进,但演化生物学家对性以及与其相关的种种方面的认识还远远不够全面。例如最近,亚马逊的茉莉鱼挑战了所有主流的性演化理论。这种小小的淡水鱼通过克隆进行生殖,但即使经过了50万代,它们的基因组质量也没有出现明显下降,而且它们在应对寄生虫和环境变化方面也做得很好。这项新的发现,以及未来更多的新发现,将继续激发科学家去挑战这一演化生物学中的女王谜题的热情。
如果我们幸运的话,演化之树上的女王也将继续让我们揭示她无穷的秘密中的星星点点。
参考资料:
[1] Vrijenhoek, R., Dawley, R., Cole, C. & Bogart, J. in Evolution and Cytology of Unisexual Vertebrates (eds Dawley, R. & Bogart, J.) 19–23 (Univ. State New York, New York, 1989).
[2] Wilson, E. O. The Diversity of Life (W. W. Norton, New York, 1992).
[3] Otto, S.P. (2009) The evolutionary enigma of sex. Am. Nat. 174, S1–S14
[4] Lehtonen, Jussi, Michael D. Jennions, and Hanna Kokko. "The many costs of sex." Trends in ecology & evolution 27.3 (2012): 172-178.
[5] Bradley, A. J., I. R. McDonald, and A. K. Lee. "Stress and mortality in a small marsupial (Antechinus stuartii, Macleay)." General and comparative endocrinology 40.2 (1980): 188-200.
[6] Packer, Craig, and Anne E. Pusey. "Adaptations of female lions to infanticide by incoming males." The American Naturalist 121.5 (1983): 716-728.
[7] Hartfield, Matthew, and Peter D. Keightley. "Current hypotheses for the evolution of sex and recombination." Integrative zoology 7.2 (2012): 192-209.
[8] Full text of the book is available on The Project Gutenberg EBook website: https://www.gutenberg.org/files/12/12-h/12-h.htm
[9] Li XY., Lehtonen J., Kokko H. (2017) Sexual Reproduction as Bet-Hedging. In: Apaloo J., Viscolani B. (eds) Advances in Dynamic and Mean Field Games. ISDG 2016. Annals of the International Society of Dynamic Games, vol 15. Birkhäuser, Cham
[10] Fairbairn, Daphne J. Odd couples: extraordinary differences between the sexes in the animal kingdom. Princeton University Press, 2013.
[11] Kothe, Erika. "Tetrapolar fungal mating types: sexes by the thousands." FEMS microbiology reviews 18.1 (1996): 65-87.
[12] Heath, D. J. "Simultaneous hermaphroditism; cost and benefit." Journal of Theoretical Biology 64.2 (1977): 363-373.
[13] Munday, Philip L., Tetsuo Kuwamura, and Frederieke J. Kroon. "Bidirectional sex change in marine fishes." Reproduction and sexuality in marine fishes: patterns and processes (2010): 241-271.
[14] Berglund, Anders. "Sex change by a polychaete: effects of social and reproductive costs." Ecology 67.4 (1986): 837-845.
[15] Warren, Wesley C., et al. "Clonal polymorphism and high heterozygosity in the celibate genome of the Amazon molly." Nature ecology & evolution 2.4 (2018): 669.