所有生物都经历了熵的增加,表现为遗传和表观遗传信息的损失。在酵母中,随着时间的推移,由于染色质修饰蛋白对DNA断裂的重新定位,导致细胞失去了它们的身份,这是酵母衰老的标志。
近期,哈佛医学院David A. Sinclair团队在Cell 在线发表题为“Loss of epigenetic information as a cause of mammalian aging”的研究论文,该研究使用一种称为“ICE”(inducible changes to the epigenome,表观基因组诱导变化)的系统,发现表观遗传信息的紊乱导致小鼠衰老,而恢复表观基因组的完整性可以逆转衰老的迹象。
总之,这些数据与衰老的信息论是一致的,该理论认为表观遗传信息的丢失是衰老的可逆原因。

生命是基于细胞机制和存储在基因组和表观基因组中的信息之间复杂的相互作用,这可以被认为是生物“硬件”和“软件”。衰老是由“硬件”故障引起的,还是由“软件”故障引起的,还是两者都造成的,目前还不清楚。在20世纪50年代,Szilard及Medawar分别独立提出衰老是由DNA损伤引起的突变导致遗传信息丢失引起的。与衰老最相关的DNA损伤类型是双链DNA断裂(DSB),以每个细胞每天10-50次的速度发生。
然而,最近,关于基因突变是否是衰老的首要驱动因素的问题被提出了。许多类型的老细胞缺乏突变,具有较高突变率的小鼠或人的品系几乎没有显示出过早衰老的证据,哺乳动物可以从旧体细胞克隆出具有正常寿命的新个体。在发育过程中,细胞的身份是由转录网络和染色质结构指定的。为了保持最佳功能,细胞必须通过保存表观遗传信息和低熵状态来保持其身份。
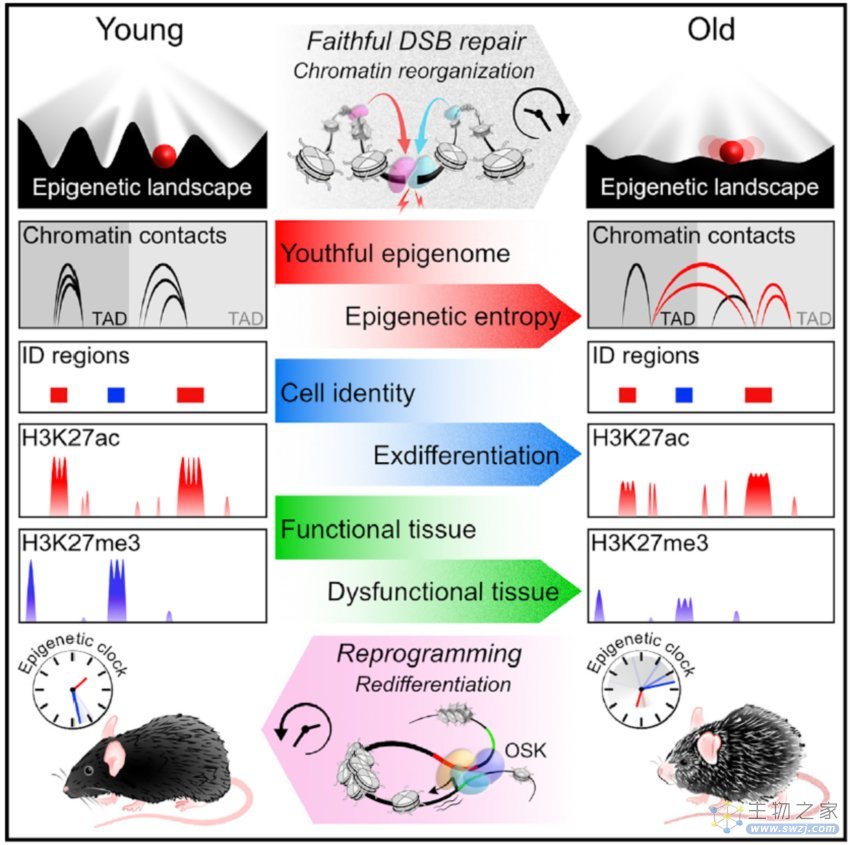
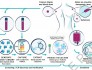
文章模式图(图源自Cell )
表观遗传信息的丢失,而不是遗传,作为衰老的潜在原因,出现在20世纪90年代的酵母研究中。沉默信息调节复合物(Sir2/3/4)从沉默交配型位点重新定位到不稳定的rDNA是不育的原因,这是酵母衰老的标志,与组蛋白占用的改变、组蛋白修饰(例如H3K56ac和H4K16ac)和基因转录一致。SIR2、组蛋白的过表达或组蛋白甲基转移酶基因SET2的缺失延长了酵母的寿命,这表明表观遗传变化不仅是一种生物标志物,而且是酵母衰老的原因。
与衰老相关的表观遗传变化,包括DNA甲基化(DNAme)模式的变化,H3K4me3, H3K9me3和H3K27me3,也可见于多细胞生物中。例子包括缺乏H3K4三甲基化复合物的蠕虫或过度表达Sir2基因的果蝇的寿命延长,以及长寿裸鼹鼠的相对稳定的表观基因组。许多表观遗传变化遵循特定的模式,包括表观遗传时钟的特定CpGs的甲基化。
哺乳动物表观基因组随时间变化的原因尚不清楚。线索还是来自酵母。酵母中的一个主要驱动因素是DSB,其修复需要表观遗传调控因子Sir2, Hst1, Rpd3, Gcn5和Esa1。染色质修饰剂的重新定位或“RCM”假说和随后的“衰老的信息论”提出,真核生物的衰老是由于转录网络和表观遗传信息随着时间的推移而丢失,由一种保守机制驱动,该机制进化为共同调节对细胞损伤的反应,如DSB或挤压损伤。
为了测试表观遗传变化是否是哺乳动物衰老的原因,该研究开发了系统来降解和重置细胞和小鼠的表观遗传信息。该研究的数据表明,哺乳动物的衰老相当于“软件”问题,是表观遗传信息损坏的结果,可以从现有的备份副本中恢复。
“该研究历经13年,我们相信我们的研究是第一个表明表观遗传变化是哺乳动物衰老的主要驱动因素的研究,”该论文的通讯作者、哈佛医学院David Sinclair说。该团队广泛的一系列实验证实了人们期待已久的事实:DNA变化不是衰老的唯一原因,甚至不是主要原因。相反,研究结果表明,染色质的化学和结构变化在不改变遗传密码本身的情况下加速衰老。
参考消息:
https://www.cell.com/cell/fulltext/S0092-8674(22)01570-7
https://medicalxpress.com/news/2023-01-loss-epigenetic-aging-reverse.html