人类基因组的破译加上生物技术的进步,特别是单克隆抗体(mAbs)的里程碑式发现,使得生物治疗极大地改善了患者的生存时间和生存状态。目前,已经有超过300种生物疗法获得美国食品和药物管理局(FDA)批准,其中许多被批准用于治疗自身免疫和炎症性疾病。
这些新药在带给人类巨大利益的同时,也让我们已经学到了很多东西,从开发这些药物的经验中得到反思。这些概念为许多此类疾病的潜在免疫致病机制提供了重要的见解,并将指导未来的药物发现和开发工作。
患者和疾病异质性
患者的不同临床表现也可能存在异质性。在银屑病和相关的银屑病关节炎(PsA)中,靶向IL-23/17通路可显著改善皮肤表现。这些药物证明了关节炎的临床改善,而相对于在皮肤中观察到的获益程度较低,这意味着影响关节炎还同时有其他炎症途径的参与。
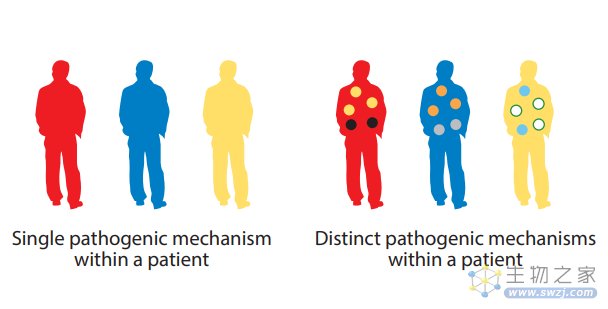
不同的病理生理机制可以驱动不同的临床表现,而选择合适的临床终点是至关重要的。这种异质性进一步表明,在某些疾病中,可能需要联合治疗才能使特定患者的临床获益最大化。
生物标志物对药物开发的成功至关重要
一种药物是否有效取决于它能否在关键试验中达到预先指定的临床终点。为了提高成功的可能性,生物标志物有助于在药物开发过程中最大限度地提高临床效益和成功的可能性。
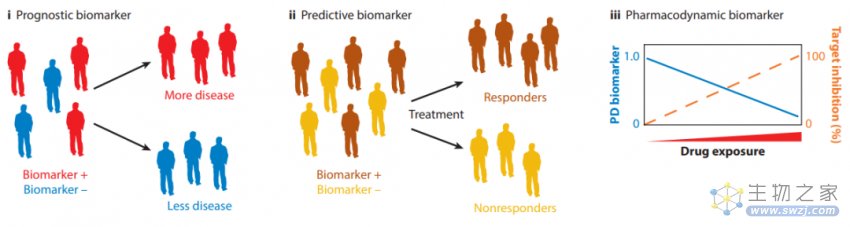
预后生物标志物识别的患者亚群具有不同的临床进程,预测性生物标志物可用于在治疗前确定更可能从治疗中获益的患者,药效生物标志物报告了药物与靶点结合的能力,提供了有关药物剂量和时间的重要信息。在药物开发中使用预测性、预后性和药效生物标志物可以缓解患者异质性的问题,丰富临床结果的测量方法,并确定是否实现了足够的靶向抑制。
遗传缺陷作为临床疗效或
安全性预测因子的局限性
单核苷酸多态性(SNPs)以及由遗传改变或人为导致获得或丧失功能(部分或完全)的等位基因,由此产生的表型分析提供了对基因功能的重要见解。然而,利用其指导药物开发在疾病适应症或安全问题上存在局限性,其中包括:
1. 疾病的发病机制在疾病的起始期和效应期存在差异。由于大多数临床试验都涉及有症状的患者,如果其致病机制不同,干扰疾病发生途径的疗法在效应期不太可能有效。
2. 在评估潜在的安全问题时,基因缺陷生物(包括人类)对感染性挑战表现出明显的敏感性。然而,在成年期出现疾病的患者由于在治疗干预前发生的环境和感染暴露或免疫接种而受益于现有的免疫力。因此,对于基因受损的人类或小鼠,抑制肿瘤坏死因子(TNF)-α、白介素(IL)-6或IL-12/23的疗法比预测的要安全得多。
3. 通常用于临床前研究的在无特定病原体(SPF)条件下饲养的动物,他们的免疫系统更像新生儿,而不是有抗原经验的成年患者,因此,在现实环境中可能错误地预测治疗干预对成人机体的生物学后果。例如,SPF饲养的小鼠的微生物群不同于生活在自然环境中的小鼠,而拥有自然微生物群的小鼠比SPF饲养的小鼠更能重现CD28超抗原和抗TNF-α单克隆抗体在人类临床试验中观察到的临床效果。
4. 最后,使基因完全失活可能是有害的,而半衰期短得多的小分子抑制剂可在不完全抑制的剂量下使用,以最大限度地提高治疗指数(即提供疗效的剂量与毒性的比率),如Janus激酶(Jak)抑制剂。
临床试验失败的经验
阴性的临床试验结果并不一定意味着所涉及的生物途径与疾病无关。临床试验失败至少有四个主要原因:
1. 药物设计:候选药物的分子特性研究不充分,包括靶向特异性、亲和力和效价、引起限制性靶向抑制不良事件的非靶向效应、药物对疾病作用部位(如大脑)的生物分布不良以及药代动力学特性差,都可能会导致阴性结果。敏感的药效生物标志物的严格应用通常可以识别这些原因。
2. 研究设计:未能为研究期间具有足够疾病活动性或疾病进展可能性的研究人群制定适当的进入标准,可能会对临床试验检测治疗效果的敏感性产生负面影响。安慰剂组需要接受伴随性标准治疗(SOC)并保持较高的依从性,如果未能解释安慰剂组的临床效果,也将减少衡量疗效的治疗窗口。最后,将适当的临床终点与靶向生物途径相匹配,对于临床表现异质的多因素疾病至关重要。
3. 患者选择:如果未能做合适的区分,疾病异质性将稀释临床试验的治疗效果。所以利用合适的生物标志物作为诊断标准至关重要,在第2阶段概念验证试验中,需要保证足够的患者数量(纳入诊断阳性和阴性患者),以便提供适当的能力,检测诊断阳性患者亚群的临床益处,以验证该诊断。在许多情况下,需要提供一个合适的界限来定义诊断阳性。
4. 靶标选择:一个设计良好的试验应该能够排除原因1-3。如果即使是这样,依然得到负面的结果,这可能意味着,尽管临床前的信息导向该适应症靶点,然而这个治疗概念实际上依然是错误的。
治疗靶点与开发药物
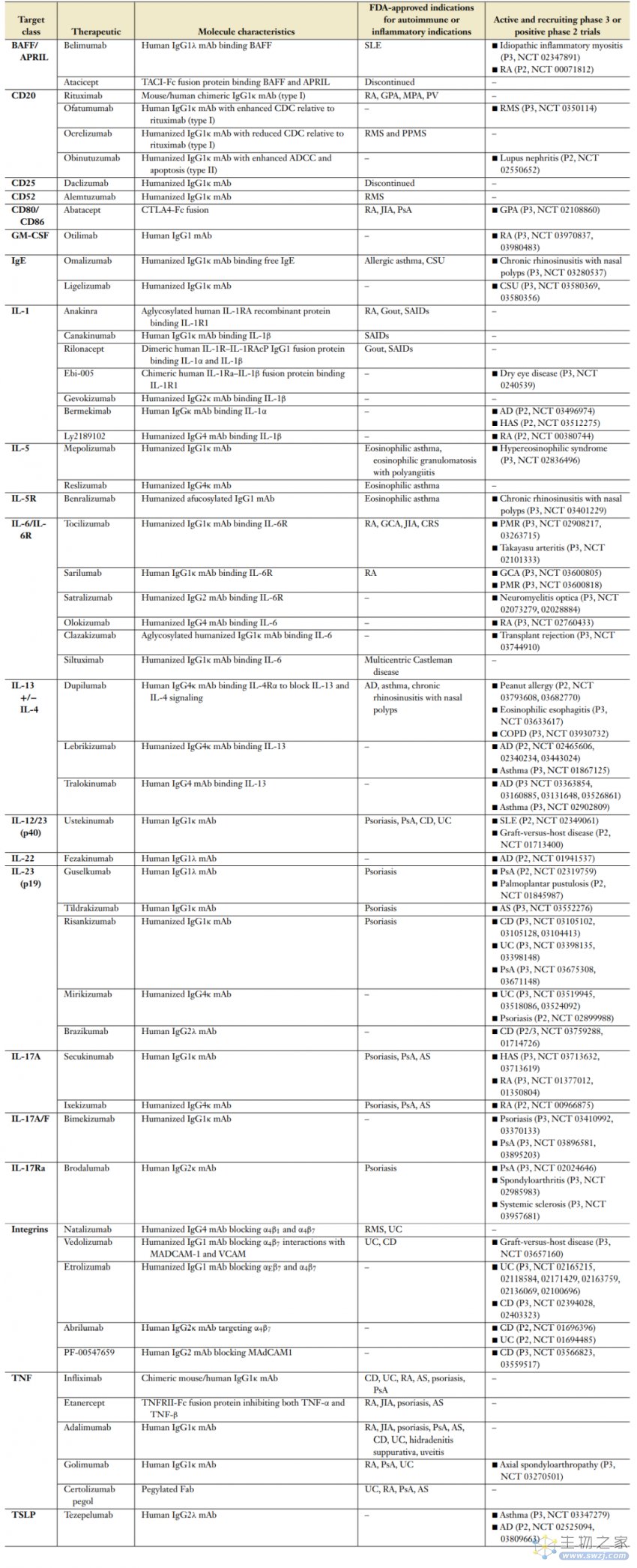
下表列出了经美国FDA批准用于自身免疫疾病适应证,以及处于关键Ⅲ期临床试验或阳性的Ⅱ期临床试验中的生物疗法。
白介素-1
IL-1α和IL-1β定位于对无菌状态下和微生物介导的炎症反应的迅速响应。当上皮细胞、内皮细胞和血小板对组织损伤作出反应时,IL-1α作为一种报警素发挥作用。相反,髓系来源的亚型IL-1β是作为一种非活性蛋白合成的,需要炎症小体驱动的蛋白水解过程才能发挥活性。IL-1α/β属于较大的IL-1细胞因子家族,它利用IL-1受体辅蛋白(IL-1RAcP)与IL-1R1亚基配对产生MyD88依赖性信号。IL-1α/β功能也受天然拮抗剂IL-1Ra、sIL-1RAcp和诱饵受体IL-1R2的调控。IL-1α/β表达失调与以发热、皮疹、关节炎和器官特异性炎症为特征的多种全身性自身炎症性疾病(SAID)有关。
美国FDA已经批准了三种不同性质的生物疗法,但都是针对IL-1的。Anakinra是一种天然拮抗剂IL-1Ra的改良版本,它同时抑制IL-1α和IL-1β。Anakinra被批准用于治疗RA和CAPS,其半衰期短,为5-6小时,因此需要每天皮下注射。Anakinra对痛风也有效,尿酸单钠晶体刺激NLRP3激活和IL-1β产生。Canakinumab是一种IL-1β中和抗体,每4-8周皮下给药一次,被批准用于治疗全身性青少年特发性关节炎(SJIA)和SAIDs。Rilonacept编码IL-1R和IL-1AcP的胞外结构域,与IgG的Fc部分相连,优先中和IL-1β,批准用于SAIDs的每周皮下给药。
白介素-6
IL-6是gp130细胞因子家族的一员,是许多稳态和炎症过程的中心组成部分。IL-6最初被鉴定为促进B细胞分化的T细胞衍生因子,现在因其在适应性免疫中的多效性活性而受到重视。它有助于Th17和T滤泡辅助细胞(Tfh)的分化,驱动髓样细胞分化,并与Th2细胞因子协同促进巨噬细胞极化为促纤维化表型。IL-6在肝脏急性期反应中起重要作用,增加包括C反应蛋白(CRP)在内的炎症蛋白,增强补体与病原体或死亡细胞的结合,对内皮细胞功能和上皮细胞完整性也有重要影响。
IL-6的广泛生物活性源于其调节多个细胞靶点的复杂性质。IL-6由多种细胞合成,并通过三种不同的细胞表面信号机制(即经典信号、反式信号和反式呈递)激活靶细胞,最终产生JAK/STAT信号。
Tocilizumab和sarilumab是FDA批准的两种IL-6R阻断性单抗,许多其他药物正在临床开发中。Tocilizumab和sarilumab被美国FDA批准用于治疗中重度RA,其中受影响关节滑液中IL-6水平升高,血清IL-6水平与疾病活动相关。这两种药物都能改善RA患者的炎症症状并减少影像学进展。
细胞因子释放综合征(CRS)是一种急性全身炎症性疾病,与许多基于抗体的治疗、化疗和T细胞参与的免疫治疗(如CAR-T细胞)以及严重感染相关。与T细胞参与疗法相关的CRS被认为是由活化的T细胞产生TNF-α,进而触发单核细胞和活化的巨噬细胞产生IL-6和IL-1β。CRS症状(例如发烧和低血压)的缓解通常在单剂量的Tocilizumab后实现。给药tocilizumab似乎并不影响T细胞参与疗法的抗肿瘤效果。
肿瘤坏死因子
TNF-α主要由免疫细胞和内皮细胞产生,并通过促炎信号和细菌产物显著上调;TNF-β(LTα)主要由淋巴细胞产生。TNF-α以三聚体膜结合形式(mTNF)表达并经历蛋白水解裂解产生可溶性三聚体sTNF。TNFR1广泛表达,而TNFR2主要表达于神经元、免疫细胞和内皮细胞。TNF-α具有多效性功能。TNF-α对病原体的最佳防御、淋巴器官的正常发育以及在神经元再髓鞘化、心脏重塑和软骨再生中的重要修复作用都是必需的。
目前有五种TNF抑制剂被美国FDA批准用于临床,全部都靶向TNF-α,其中依那西普另外抑制TNF-β。肿瘤坏死因子拮抗剂的分子类型反映了近二十年来生物治疗药物的发展。依那西普(TNFR2-Fc)是美国FDA批准的第一个Fc融合蛋白;infliximab是第一代嵌合单抗;adalimumab是来源于噬菌体展示的人源单抗;golimumab是来源于表达人IgG的转基因小鼠的人源单抗;certolizumab-pegol是一种从小鼠杂交瘤中分离的人源化Fab片段,经聚乙二醇化可延长其体内半衰期。
大多数肿瘤坏死因子拮抗剂显示出共同的临床疗效。然而,依那西普在克罗恩病(CD)的随机双盲安慰剂对照试验中无效,可能反映了TNF抑制剂治疗机制的差异。除了中和sTNF-α外,抗TNF抗体和certolizumab-pegol(而不是依那西普)可通过结合mTNF或阻断抗凋亡信号诱导固有层T细胞凋亡。此外,抗TNF单抗(而非依那西普或certolizumab-pegol)可通过Fc依赖机制诱导M2型创伤愈合巨噬细胞应答。因此,TNF拮抗剂之间的机制差异可能是导致其在CD中不同临床效果的原因。
CD20
CD20是一种B细胞标志物,其表达始于早期的前B细胞,但在向浆细胞的终末分化过程中丢失。它是一种四肽蛋白,是B细胞发挥最佳功能和免疫反应所必需的。
Rituximab是一种嵌合单抗,是美国FDA批准的第一种治疗B细胞非霍奇金淋巴瘤的抗CD20单抗。基于其在肿瘤学中令人惊讶的良好安全性,研究人员开始探索其在严重自身免疫性疾病患者中的应用。Rituximab目前被美国FDA批准用于治疗多种自身免疫性疾病。由于CD20既不在HSC上表达,也不在终末分化的浆细胞上表达,因此抗CD20单抗选择性靶向CD20+B细胞与先天性B细胞缺乏具有不同的免疫学后果。例如,用抗CD20抗体治疗后,血清IgG水平没有显著影响,而严重的X连锁无丙种球蛋白血症(其中B细胞不发育)则观察到缺乏免疫球蛋白。
最近的一项Ⅱ期研究使用了obinutuzumab,一种具有增强ADCC和诱导B细胞凋亡能力的II型抗CD20单抗,已报道狼疮性肾炎患者的临床获益(NCT 02550652)。需要对obinutuzumab进行进一步的研究,以确认额外的B细胞消耗是否对狼疮性肾炎有益。
BAFF/APRIL肿瘤坏死因子超家族成员
BAFF和APRIL是TNF配体超家族的两种II型跨膜蛋白,它们的受体BAFFR、BCMA和TACI在B系细胞的存活和成熟以及功能中起着重要作用。
血清BAFF和APRIL水平在许多自身免疫性疾病中升高,包括SLE、Sjögren综合征和RA。Belimumab是一种人IgG1λ单抗,被批准用于治疗活动性自身抗体阳性的SLE。它抑制sBAFF三聚体,但不与mBAFF或APRIL结合。在关键的临床试验中,与SOC组相比,belimumab在SLE应答指数和健康相关生活质量终点方面有更大的改善。此外,belimumab降低了循环中原始B细胞、活化B细胞和浆细胞的数量;抗dsDNA水平降低,补体水平正常化。与对循环记忆B和T细胞缺乏影响一致,治疗一年后,先前存在的抗肺炎、破伤风和甲型流感抗体没有受到影响。
Atacicept是融合TACI胞外区的Fc融合蛋白,结合BAFF、APRIL和BAFF/APRIL异源聚体。在系统性红斑狼疮患者中应用Atacicept治疗后,血清IgM(∼70%)、IgG(∼30–40%)和IgA(∼50–60%)迅速下降,抗双链DNA抗体下降了∼40%。终止治疗后,抗体和自身抗体水平恢复到预处理水平。
整合素家族
整合素通过调节白细胞与血管的粘附来调节免疫细胞的运输,并促进白细胞向组织中的外渗。整合素异二聚体由18个α亚单位和8个β亚单位之间的配对组成,形成24种不同的受体复合物,它们在表达模式、配体特异性和功能上存在差异。
在MS中,α4β1(CD49d/CD29、VLA4)在T和B细胞上表达,并通过VCAM1与血脑屏障内皮细胞的结合促进淋巴细胞向中枢神经系统(CNS)的外渗。在胃肠道中,表达于T细胞上的α4β7可与Peyer氏斑高内皮小静脉和固有层小静脉上的MADCAM-1结合,促进炎症性肠病(IBD)致病性T细胞的外渗。
Natalizumab是一种α4阻断性单抗,被批准用于治疗CD和RMS患者。然而,RMS抑制淋巴细胞进入中枢神经系统的有效性的机制基础也为进展性多灶性白质脑病(PML)的罕见病例的发展提供了基础。第二代抗体,包括vedolizumab(对α4β7复合物有选择性)和etrolizumab(抗β7),通过针对IBD的肠道特异性整合素降低PML的可能性。Vedolizumab可诱导临床反应和缓解,美国FDA已批准用于溃疡性结肠炎(UC)和CD。除了阻断α4β7:MADCAM1相互作用外,etrolizumab还干扰E-钙粘蛋白+肠上皮细胞对αEβ7+IELs的保留。在一项Ⅱ期临床试验中,与对照组相比,对UC患者使用etrolizumab治疗导致临床病情缓解的可能性更大。由于αEβ7+IELs是UC患者炎症细胞因子的重要贡献者,因此阻断α4β7和αEβ7整合素可能有额外的临床益处。
抗αL抗体Efalizumab阻断T细胞和B细胞表达的αLβ2(LFA1)与ICAM1的相互作用。除了阻断整合素外,Efalizumab还下调LFA-1的表达,LFA-1是T细胞与抗原呈递细胞(APC)形成突触所必需的。Efalizumab被美国FDA批准用于治疗中度至重度斑块型银屑病。然而,4名使用Efalizumab治疗3年以上的患者出现了PML,该药物已退出市场。
IL-12和IL-23及其下游效应物
IL-12和IL-23是两种密切相关的APC衍生的异二聚体细胞因子,分别由p35和p19亚单位组成,它们与一个共同的p40亚单位配对。这两种细胞因子,主要的应答细胞是T细胞和固有淋巴细胞(ILCs),在不同的细胞类型中,IL-12上调Th1主转录因子T-bet的表达并诱导IFN-γ的产生。以类似的方式,IL-23刺激诱导转录因子RORγt的表达和下游细胞因子IL-17A、IL-17F、IL-22和GM-CSF的表达。疾病的临床前模型和全基因组关联研究表明IL-12/23通路参与炎症性肠病和银屑病。
设计用于干扰IL-12/23通路各种成分的多种疗法已进入临床。两种单抗(ustekinumab和briakinumab)靶向p40亚单位,从而中和IL-12和IL-23。最近,效应细胞因子的IL-17家族引起了广泛关注,有5种单抗选择性中和IL-17A;2种单抗中和IL-17A和F;brodalumab靶向IL-17RA,一种IL-17A、B、C、E和F成员的共同受体。然而,虽然IL-17B-E的不同功能已被描述,但这些细胞因子不受IL-23的调节,它们在人类疾病中的作用仍未被充分研究。
此外,对阻断IL-22(fezakinumab)、IFN-γ(fontolizumab)和GM-CSF(namilumab和mavrilimumab)治疗的研究将提供进一步的见解,尽管它们还处于早期研究。
机遇与挑战
在过去的三十多年里,生物疗法的进展为患者带来了巨大的益处,并对基础免疫学和疾病发病机制有了重要的认识。这些临床实验也揭示了新疗法发展的重大挑战,部分原因是疾病异质性和临床研究设计,包括患者选择、终点选择以及生物标志物的发现和研究。而新药研发工作仍有大量未满足的临床需求,发现新靶点和新生物标志物的工具和方法不断涌现,其中一些令人兴奋的进展包括:
1. 基于人类遗传学的靶点发现:DNA测序成本的降低使得人类遗传学在靶点发现方面有了新的应用,改变疾病严重程度的遗传因素的修饰筛选可能为治疗靶点提供线索。例如,IL-6R被确定为阿尔茨海默病发病年龄的候选调节因子。
2. 新型药物平台:尽管迄今为止大多数生物疗法都集中于中和单个靶点,但生物疗法的进展现在允许在单个分子内靶向多种不同的抗原或途径。随着基因编辑技术的出现,将功能设计到人类细胞中也成为可能。在癌症中,CAR-T细胞已显示出良好的应用前景,而在自身免疫中,CAR-Treg细胞正被用作诱导抗原特异性耐受自身抗原的手段。随着CRISPR技术的迅速发展,细胞可以被设计成执行多种功能,而这种工作的复杂程度几乎没有理论上的限制。因此,工程细胞疗法具有巨大的前景。
3. 微生物群:在过去的十年里,越来越明显的是,生活在我们体内的微生物在体内平衡和疾病中扮演着至关重要的角色。患者的微生物组可以作为重要的预后或预测疾病的生物标志物。目前正在进行多种针对微生物组的努力,以追求治疗效果,这些技术在粘膜疾病领域具有特别的前景。
4. 了解干预后的生物反应:随着新技术和技术平台敏感性的提高,我们通过DNA、表观遗传学、单细胞转录组学、代谢组学、蛋白质组学、外显子体和成像来研究人类生物学的能力将为人类生物学和治疗干预后的反应提供更多的见解。对应答者和无应答者之间的差异进行分析可能会揭示更多的靶向途径。
5. 大数据的应用:随着临床、生物标志物和应答数据的积累和适当的注释,将机器学习应用于这些数据集可能会提供新的治疗途径。
理解人类疾病生物学和开发新疗法的许多挑战被这些新兴的新技术和见解所平衡。它们的应用无疑将增进我们对人类疾病的了解,并提高我们为尚未满足的医疗需求提供临床益处的能力。
参考文献:
1.30 Years of Biotherapeutics Development-What Have We Learned? Annu Rev Immunol. 2020 Apr 26;38:249-287